



Bistability and Hysteresis in Biological Systems
Bistability and Hysteresis in Biological Systems: From Mathematical Models to Cellular Behavior


Biological systems are remarkably adept at processing information and making decisions. Unlike the binary components of computers, living cells must reliably interpret noisy signals in fluctuating environments. One of the most fascinating mechanisms enabling this robustness is bistability, which refers —a property where a system can exist in one of two stable states depending on external conditions. This phenomenon provides cells with a form of memory and allows for sharp, switch-like responses to gradually changing stimuli.

At its core, bistability emerges when a system possesses two stable steady states separated by an unstable equilibrium point. Conceptually, you can envision this as a ball resting in one of two valleys divided by a hill, as depicted above. The ball remains stable at the bottom of either valley but requires a sufficient push to overcome the hill and transition to the other valley.
In mathematical terms, bistable systems typically arise from nonlinear interactions, often involving positive feedback loops. A classic example is the auto-activation of a protein that promotes its own production. When properly tuned, such a system can maintain either low or high protein levels as stable states.
🧬 T7 RNA Polymerase: A Biological Case Study
A compelling real-world example of bistability comes from studies of mutant T7 RNA polymerase (T7R) expression systems. Researchers observed natural bistability in this system and developed a mathematical model combining two positive activation cycles—one representing auto-activation and another capturing metabolic costs, growth retardation, and dilution effects. This model was qualitatively similar to actual cellular responses, and thus the following equation was developed to represent the system:
The first term in the equation above describes T7R production (including self-activation), the second term represents utilization for growth, metabolic costs, and dilution, and the third term accounts for intrinsic decay.
With parameter values δ=0.01, α=10, Φ=20, and γ=10, this system exhibits bistability. To find a steady state of this system we set the differential equation to zero and then plot the product term against the two degredation terms to see where they intersect — the result is two stable steady states that occur at a very low and very high value of T7R, and an unstable steady state in the middle. The code below generates plots to show where these stable states are:
import numpy as np
import matplotlib.pyplot as plt
# Input arameters
delta, alpha, phi, gamma = 0.01, 10, 20, 10
# Define the production and degradation terms
def production(T7R):
return (delta + alpha * T7R) / (1 + T7R)
def degradation(T7R):
return (phi * T7R) / (1 + gamma * T7R) + T7R
# Different T7R ranges for subplots
T7R_ranges = [np.linspace(1, 100, 100), np.linspace(0.0001, 0.01, 100), np.linspace(0.01, 1, 100), np.linspace(1, 10, 100)]
# Plotting
fig, axs = plt.subplots(2, 2, figsize=(12, 10))
fig.suptitle("Bistability in T7 RNA Polymerase Expression (Various Ranges)")
# Plot each range in a separate subplot
for i, T7R_vals in enumerate(T7R_ranges):
row, col = divmod(i, 2)
prod_vals = production(T7R_vals)
deg_vals = degradation(T7R_vals)
axs[row, col].plot(T7R_vals, prod_vals, label="Production")
axs[row, col].plot(T7R_vals, deg_vals, label="Degradation")
axs[row, col].set_xlabel("T7R")
axs[row, col].set_ylabel("Rate")
axs[row, col].legend()
axs[row, col].grid(True)
axs[row, col].set_title(f"T7R range: [{T7R_vals[0]}, {T7R_vals[-1]}]")
plt.tight_layout(rect=[0, 0, 1, 0.95])
plt.show()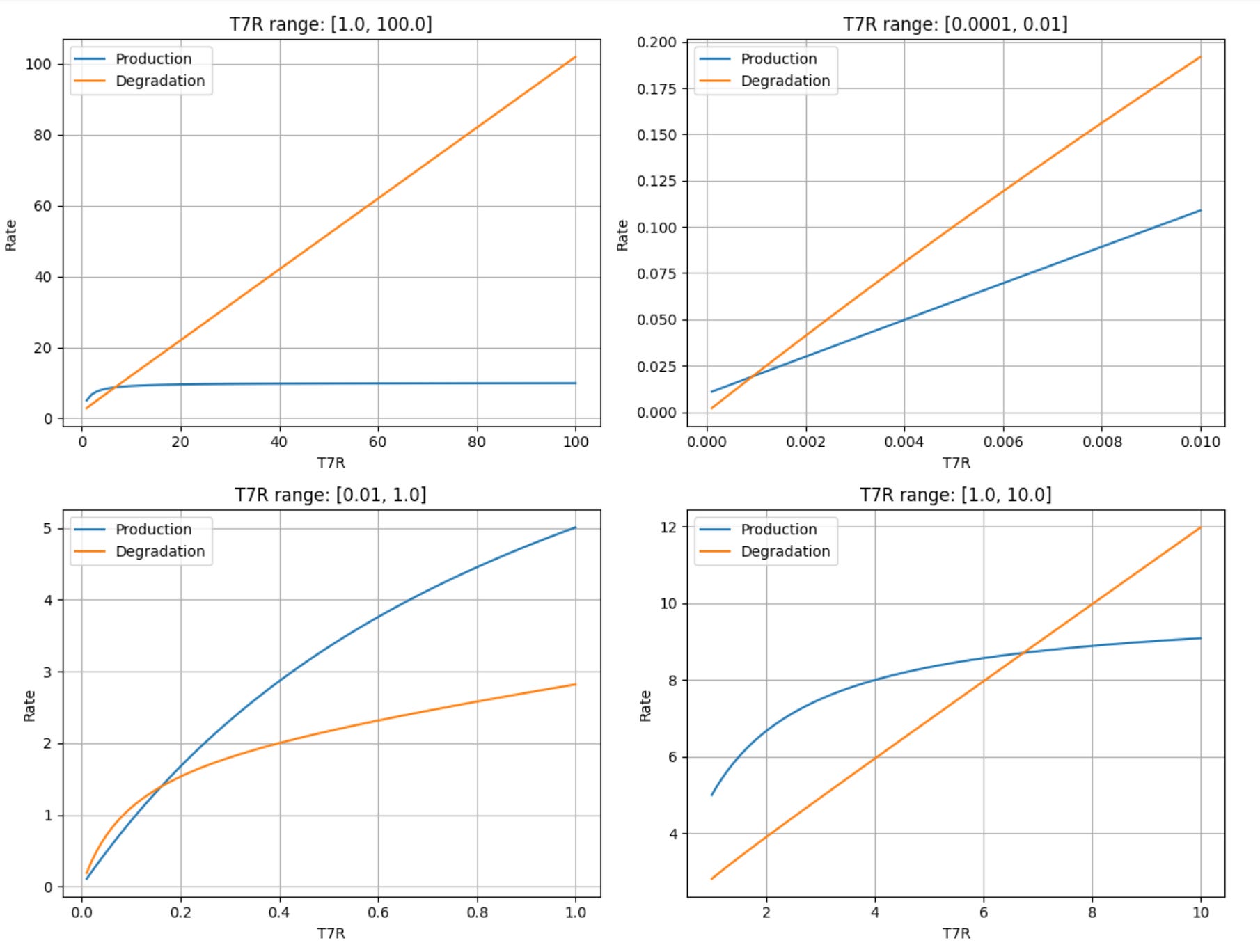
🧬 From Bistability to Hysteresis: The Two-Gene System
Moving beyond single-variable systems, let's consider a slightly more complex two-gene network where genes X and Y regulate each other, with an external signal S influencing the system, as depicted below:

The mathematical representation of this system is as follows:
What makes this system particularly interesting is that it demonstrates hysteresis—a form of cellular memory where the response depends not only on the current signal value but also on the system's history. This occurs because the two stable states are separated by an energy barrier that the system must overcome to transition between states.
To observe hysteresis experimentally, one would gradually increase signal S from a low value, allowing the system to reach steady state after each increment. At some critical threshold, the system suddenly jumps to a higher X state. Then, starting from this high state and gradually decreasing S, the system maintains its high state until S drops significantly below the original activation threshold.
🧬 Biological Significance and Applications
The bistability and hysteresis properties have profound implications for cellular decision-making. For examplem, they enable:
Noise filtering: Small fluctuations around a steady state don't change the system's behavior, preventing unecessary (or counter productive) responses to transient signals.
Commitment to developmental fates: Once a cell commits to a particular fate, it stays committed even if the initiating signal diminishes.
All-or-none responses: Cells can make discrete decisions rather than exhibiting graded responses to continuous signals.
Bistable switches appear throughout biology, from the lac operon in bacteria to the cell cycle control system in eukaryotes. Additionally, the genetic toggle switch, represents a synthetic biology implementation of bistability that has become a foundational tool in the field.
Additionally, coupling bistable switches with other network motifs like oscillators can generate complex behaviors such as excitability and pulsatile responses.
Understanding these dynamics is crucial for both deciphering natural biological processes and engineering synthetic circuits with predictable behaviors. As biotechnology advances, the principles of bistability and hysteresis serve as essential design tools for creating cellular systems with memory, decision-making capabilities, and robust responses to environmental changes.
One critical remark/addition - these dynamic behaviors have been amply studied in ecological theory with 2 and 3 species systems since Robert May's work in the 1970s. Some of these behaviours have been incorrectly characterised in other literature as "tipping points".
I believe that the inverse of this problem will be extremely valuable if solvable. By that I mean, given the output data of some biological system, and no prior knowledge of the underlying dynamics, can you infer the existence and properties of certain steady states in the system?
I wonder if many disease processes are well-described by changes in the underlying dynamics of a governing system, and if therapeutic interventions are better thought of as inputs into a system, rather than a patch to what is broken.